
超高分文章!有关环状RNA抑制结直肠癌进展新成果
近日,南方医科大学基础医学院病理学系李学农教授团队在国际权威期刊《Molecular Cancer》(中科院SCI分区小类1区,影响因子41.44)上刊发了题为“circEXOC6B interacting with RRAGB, an mTORC1 activator, inhibits the progression of colorectal cancer by antagonizing the HIF1A RRAGB mTORC1 positive feedback loop”的研究论文。该文作者通过高通量测序筛选及qPCR等技术,发现了在结直肠癌显著下调的环状RNA circEXOC6B,阐明了环状RNA circEXOC6B与RRAGB结合抑制其异源二聚体的形成,破坏其与RRAGC /D的相互作用,从而阻断mTORC1通路对营养的感知,抑制HIF1A-RRAGB-mTORC1正反馈环,使肿瘤细胞生长受到抑制并增强常用化疗药物5-FU诱导的凋亡。
研究背景
结直肠癌是全球发病率第二,死亡率排名第三的恶性肿瘤,常用的放化疗治疗方法产生的耐药性严重限制了治疗效果,探索结直肠癌发生发展的分子机制是改善结直肠癌患者预后的关键。近年来,越来越多的研究表明,环状RNA在调节肿瘤发展和化疗耐药性中起着至关重要的作用。使用高通量RNA测序,作者先前发现circEXOC6B在结肠癌中表达下调。然而,其在结直肠癌(CRC)中的作用和机制仍然未知。
研究方法
通过qRT-PCR实验检测CRC组织中circEXOC6B的表达,进行体内和体外功能实验以确定circEXOC6B对CRC发展的抑制作用,通过RNA-pull down、质谱、co-IP、荧光原位杂交和免疫荧光,研究circEXOC6B与CRC生长和5-氟尿嘧啶诱导的细胞凋亡的可能机制。进行染色质免疫沉淀、双荧光素酶测定、Western Blot和免疫组化实验用于探讨HIF1A调节RRAGB转录的机制。
研究结果
1. circEXOC6B在CRC组织中表达下调
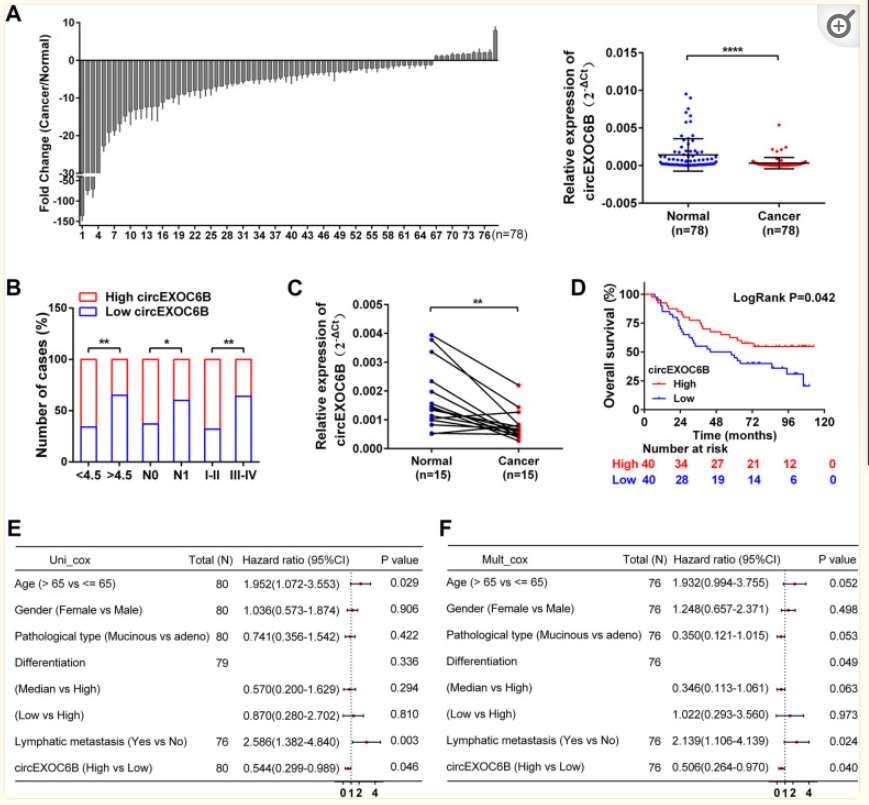
作者首先通过qRT-PCR实验测定CRC组织中circEXOC6B的表达,如图1A所示,与相邻的非癌组织(正常)相比,在78例CRC组织(癌症)中,circEXOC6B表达下调。为了探索circEXOC6B表达与CRC患者的临床病理参数之间的关系,再将78例CRC患者分为高表达组和低表达组(n= 39),根据circEXOC6B的中值表达进行统计分析,表明circEXOC6B的表达与肿瘤大小、淋巴转移和TNM分期呈负相关(图1B和表S3)。此外,还检测了结肠癌cDNA芯片中circEXOC6B的表达,该芯片包含80例癌组织和15例癌旁组织。qRT-PCR结果显示,circEXOC6B在癌组织中的表达也低于匹配的相邻(正常)组织(图1C)。Kaplan-Meier生存分析显示,circEXOC6B高表达患者的总生存率(OS)(54.8%)高于circEXOC6B低表达患者总生存率(OS)(20.6%)(P= 0.042)(图1D)。circEXOC6B表达较高的患者死亡风险较低(图1E)。最后,多元回归分析进一步证实,高circEXOC6B表达可将死亡风险降低49.4%(图1F)。这些结果表明,circEXOC6B的表达与与CRC患者总生存期相关。
2. 体外实验和在体实验表明,circEXOC6B可抑制CRC细胞的生长
为了研究circEXOC6B在CRC进展中的作用,作者使用特异性靶向circEXOC6B的siRNA敲低了CRC细胞中circEXOC6B的表达,并使用circEXOC6B过表达载体上调了circEX0C6B的基因表达(图2A和S2A)。流式细胞术和CCK8、EdU和集落形成实验表明,敲低circEXOC6B的表达促进CRC细胞的增殖、克隆形成和细胞周期,而过表达circEXOC6B则抑制癌细胞生存能力和克隆形成(图2B–D和S2B–E)。此外,流式实验结果表明,敲低circEXOC6B抑制了5-FU诱导的CRC细胞凋亡,而过表达circEXOC6B浓度具有相反的作用(图2E)。

3. circEXOC6B与RRAGB结合,干扰其异二聚体的形成
为了探索circEXOC6B调控CRC进展的分子机制,首先使用FISH、PCR和qRT-PCR检测了circEXOC6B在CRC细胞系和CRC临床样本中的亚细胞定位。结果表明,circEXOC6B主要存在于CRC细胞的细胞质中(图3A和S4)。细胞质中的circRNAs可以与miRNA或蛋白质相互作用,它们与蛋白质的结合被认为是它们的主要功能。为了鉴定与circEXOC6B相互作用的蛋白质,作者在SW620和HCT116细胞中进行RNA-pull down,随后,通过质谱分析鉴定了17种蛋白质。其中,RRAGB是与circEXOC6B结合的候选蛋白。通过RNA-pull down实验,再进行western blot实验,证实了circEXOC6B和RRAGB的结合(图3C)。此外,RIP实验验证了RRAGB可以富集circEXOC6B(图3D)。为了进一步证实circEXOC6B和RRAGB的相互作用,在CRC细胞中使用FISH和IF标记circEXOC6B和RLAGB。结果显示,在CRC细胞的细胞质中,circEXOC6B和RRAGB共定位。此外,使用catRAPID数据库,预测了RRAGB与circEXOC6B的可能结合位点。结果表明,RRAGB中circEXOC6B结合的最可能区域位于250 aa位点附近(图S6)。因此,作者构建了不同长度的RRAGB表达载体(图3F)。RIP分析显示,RRAGB的223–346 aa区域与circEXOC6B的相互作用(图3G)。有趣的是,该区域包含RRAGB与RRAGC/D异二聚体的结合位点(图3H)。因此,作者推测,circEXOC6B可能会竞争性地干扰RRAGB–RRAGC/D异二聚体的形成。为了验证这一假设,用标记RRAGB表达载体转染CRC细胞后,对其进行了Co-IP测定。Western blot结果显示,RRAGC和RRAGD与RRAGB的结合随着circEXOC6B的过度表达而降低(图3I)。这些结果表明,circEXOC6B可以与RRAGB结合,并抑制RRAGB和RRAGC/D的异二聚体形成。

4. circEXOC6B抑制mTORC1通路
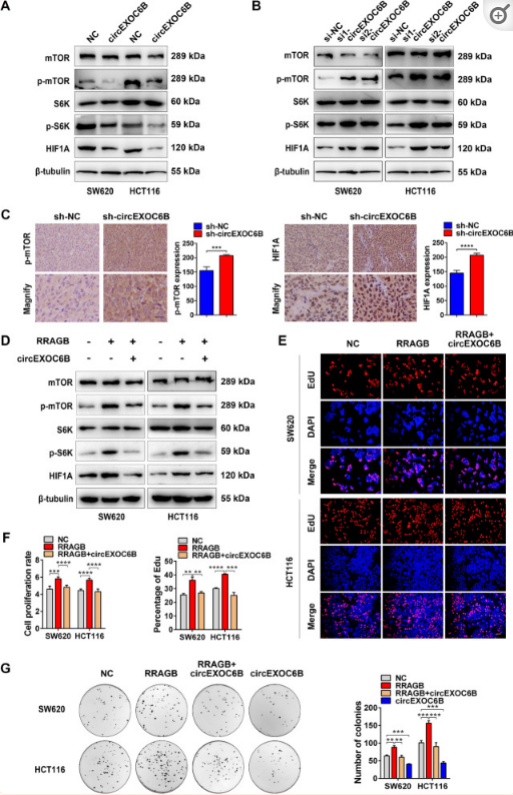
鉴于激活mTORC1途径需要RRAGB-RAGC/D异二聚体的形成,我们推断,circEXOC6B可能通过干扰异二聚物的形成来抑制mTORC1通路。Western blot结果显示,circEXOC6B的过表达降低了p-mTOR、p-S6K和HIF1A的表达,而其下调增加了它们的表达(图4A和B)。与Western Blot结果一致,IHC表明皮下肿瘤中p-mTOR和HIF1A表达也上调,circEXOC6B表达稳定沉默(图4C)。此外,作者进行了挽救试验,以进一步验证RRAGB参与mTORC1通路中circEXOC6B的调节。WesternBlot实验显示,RRAGB引起的p-mTOR、p-S6K和HIF1A升高可以通过与circEXOC6B共转染来逆转(图4D)。RRAGB诱导的CRC细胞的增殖和克隆形成性增强也可以通过circEXOC6B的过度表达来挽救(图4E-G)。这些结果支持以下假设:circEXOC6B通过竞争性结合RRAGB抑制mTORC1途径以促进CRC生长。
5. CRC具有HIF1A-RAGB-mTORC1正反馈回路,该回路被circEXOC6B拮抗
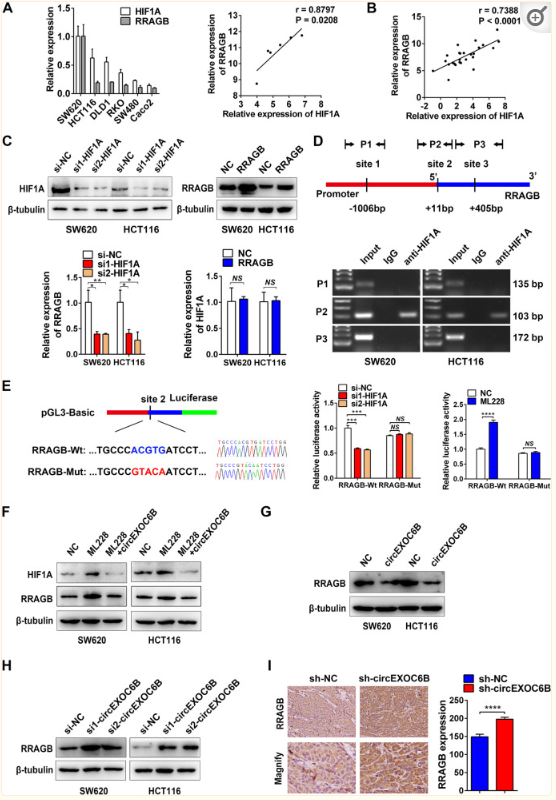
作者使用qRT-PCR实验检测六种CRC细胞系中HIF1A和RRAGB的表达时,惊奇地发现HIF1A与RRAGB表达之间存在正相关(图5B)。此外,HIF1A和RRAGB的表达水平在26个CRC组织中也呈正相关(图5A)。同时使用GEO(图S7A)和癌症基因组图谱(TCGA)(图S7B)数据集进一步验证了它们在CRC中的正相关性。这些发现促使作者去探索HIF1A和RRAGB表达之间是否存在转录调控。qRT-PCR实验结果显示HIF1A的敲低导致RRAGB mRNA表达的下调,而RRAGB的过表达对HIF1A mRNA表达没有影响(图5C)。同时作者也发现RRAGB的启动子区包含三个HRE(RCGTG),它们是转录因子HIF1A的结合位点(图5C)。随后的ChIP实验表明HIF1A与启动子的第二个HRE位点结合(图5D)。随后,作者构建了RRAGBWt和RRAGBMut(突变HRE)荧光素酶报告基因载体,进行双荧光素素酶报告基因实验(图5E)。结果表明,敲低HIF1A表达后,荧光素酶活性降低,但使用HIF1A激活剂ML228过度表达HIF1A后,荧光素酶活力增加(图5E)。这些结果表明HIF1A可以与RRAGB的启动子结合并促进RRAGB转录。mTORC1通路促进HIF1A的翻译。因此,HIF1A-RAGB-mTORC1正反馈环存在于CRC中,以促进肿瘤进展,而circEXOC6B可能通过与RRAGB结合来拮抗该环。作者使用HIF1A激活剂ML228上调HIF1A,Western Blot结果显示RRAGB水平升高。然而,circEXOC6B能够消除该效应并降低RRAGB和HIF1A的表达(图5F)。此外,circEXOC6B的过表达下调了RRAGB的表达,而circEXOC6B的敲低则提高了RRAGG的表达(图5G和H)。与Western Blot结果一致,IHC结果显示circEXOC6B表达稳定沉默的皮下肿瘤比对照组具有更高的RRAGB表达(图5I)。为了进一步验证circEXOC6B在正反馈回路中的抑制作用,使用IHC评估了22个CRC临床样本中HIF1A、RRAGB和p-mTOR的表达,包括11例circEXOC6B低表达和11例circEXOC6B高表达。结果表明,HIF1A、RRAGB和p-mTOR水平在高circEXOC6B组中低于低circEXOC6B组(图S8)。
6.circEXOC6B增强CRC细胞对5-FU的敏感性
5-FU被广泛用作CRC的一线化疗药物。然而,5-FU的有效性因固有或获得性化疗耐药性而降低。近年来,mTORC1和HIF1A通路的激活被认为是5-FU耐药的关键原因。之前的流式实验数据如图2E所示,表明circEXOC6B促进了5-FU诱导的CRC细胞凋亡。此外,发现5-FU敏感细胞系RKO、LoVo、HCT116和HT-29中circEXOC6B的表达高于5-FU非敏感细胞系SW620(癌症数据库中药物敏感性基因组:https://www.cancerrxgene.org/)(图S9)。因此,circEXOC6B可能通过抑制HIF1A-RAGB-mTORC1正反馈回路来增强CRC细胞对5-FU治疗的敏感性。为了进一步评估circEXOC6B对5-FU治疗的影响,进行TUNEL检测5-FU诱导的细胞凋亡实验。结果表明,较低的circEXOC6B表达减少了5-FU诱导的CRC细胞凋亡,而较高的circEXOC6B的表达使CRC细胞对5-FU治疗更敏感(图6A)。此外,circEXOC6B的敲低降低了凋亡标志物裂解caspase-3和裂解PARP的表达,而其过度表达增加了其表达(图6B)。与凋亡结果一致,CCK8测定显示circEXOC6B的下调拮抗了5-FU在CRC细胞增殖中的抑制作用。相反,circEXOC6B的上调增强了5-FU对增殖的影响(图6C)。此外,通过构建裸鼠成瘤,以测试circEXOC6B在体内5-FU治疗中的作用。腹腔注射5-FU后,sh-circEXOC6B组的肿瘤体积与未治疗组无差异。相比之下,5-FU治疗后,sh-NC组的肿瘤大小显著减小(图6D和E)。此外,在sh-circEXOC6B组中,凋亡标志物裂解的caspase-3的表达没有增加,但在sh-NC组中显著升高(图6F)。与caspase-3的表达一致,TUNEL结果显示,5-FU治疗后,sh-circEXOC6B组的凋亡没有变化,而5-FU处理后,sh-NC组的凋亡显著增加(图6G)。这些发现表明,circEXOC6B增强了CRC细胞对5-FU的敏感性。

结论
本研究揭示了HIF1A-RRAGB-mTORC1正反馈回路的存在,该回路驱动结直肠癌的恶性演进, 而circEXOC6B抑制结直肠癌的生长并增加5-FU诱导的结直肠癌细胞凋亡(附图)。这些结果为HIF1A和mTORC1通路之间的串扰提供了新的见解,并提出了阻断结直肠癌进展的潜在治疗靶点,揭示了环状RNA调控结直肠癌生长及化疗敏感性的新机制,为寻求结直肠癌治疗新靶点提供了新的技术途径。


